Missing something? Perhaps. Earth may once have had rings. Photo: NASA, The Blue Marble.
When I was a wee lad with a growing interest in astronomy and science fiction, I sometimes wondered what it would have been like if Earth, like Saturn, had a ring.
As it turns out, not great, apparently.
In a paper published in Science Direct, Andrew Tomkins, Erin Martin and Peter Cawood suggest Earth may indeed have had a ring during the Ordovician, around 466 million years ago (Ma) with intriguing but catastrophic consequences.
As Tomkins explains in an article for The Conversation, the first clue for the team was an anomalous series of 21 impact craters from around that time, combined with ‘deposits of limestone across Europe, Russia and China containing very high levels of debris from a certain type of meteorite.’
The team then mapped where the craters would have been 466 Ma by using models of tectonic plate movement. Turns out they were all along the equator, with none near the poles.
As Tomkins writes, ‘… we measured how much of Earth’s land surface suitable for preserving a crater was near the equator at that time. Only about 30% of the suitable land was close to the equator, with 70% at higher latitudes.’
So, odds on something peculiar was going on.
The team speculates that Earth captured a passing asteroid which got close enough to exceed the Roche limit (where a smaller body’s internal gravity is overwhelmed by the tidal forces of a larger body). The broken remains of this asteroid then formed a ring around the Earth’s equator, and over time bits of it crashed into the Earth’s surface, forming the impact craters.
Finally, Tomkins et al speculated that the ring would have cast a shadow across large swathes of the Earth’s surface, which together with atmospheric dust from the impacts may explain the sudden plunge into what’s called the Hirnantian Ice Age about 445 Ma.
The team then further speculates that this rapid cooling would have forced animals to adapt to changing conditions, leading to the Great Ordovician Biodiversification Event. Uncomfortably abbreviated to GOBE, this was when Palaeozoic fauna replaced Cambrian fauna, leading to ‘… a spectacular increase in marine biodiversity at all taxonomic levels … ‘[i]
So wearing a ring was not all bad news, I suppose.
[i] Servais, Thomas, et al ‘The Great Ordovician Biodiversification Event (GOBE): The palaeoecological dimension’, ScienceDirect. See here.
In 1977 two American microbiologists discovered a whole new branch of life: the archaea. Single cells without a nucleus, archaea are prokaryotes that for a long time were considered to be bacteria. The work of Carl Woese and George E. Fox, however, revealed that archaea are in some respects closer to eukaryotes (cells with a nucleus, which includes all multicellular life such as you and me) than bacteria.
As a result of their discovery, the traditional taxonomic tree, starting with kingdom and ending with species, was – so to speak – recapped. A new level was placed at the top of the tree – the domain – with three members: bacteria, archaea and eukaryotes.
‘So where do viruses fit in?’ I hear you ask.
Good question. Unfortunately, there isn’t a good answer. The issue is confusing because there is no clear understanding of their evolutionary development, or even if they share a common ancestor. Some may have developed from small, free-floating bits of DNA called plasmids, while others may have evolved from bacteria. It’s generally easier to hedge around the issue of whether or not viruses are actually alive rather than confronting it, and they are often simply called infectious agents or biological entities. The main sticking point is that viruses cannot replicate without infecting a host cell, and are therefore entirely dependent for their existence on ‘biological entities’ that are undoubtedly living. An example of how contentious this argument can be, however, is that the bacteria that causes the STD chlamydia can only exist in a host cell … therefore, should this bacteria be considered technically alive?
Now, to complicate matters even further, a preprint article published in bioRxiv earlier this year announced the discovery of ‘viroid-like colonists’ the authors call ‘Obelisks’, which sounds awfully like the proposed title for a 1950s sf monster movie set somewhere in Death Valley.
However, Obelisks are minute, not monstrous. In fact, they’re small enough to fit inside bacteria. The article’s first sentence describes them as a ‘previously unrecognised class of viroid-like elements that we first identified in human gut metatransciptomic data.’ Basically, they were found in samples of human poo. I think I’d prefer to research giant creatures rampaging through Death Valley, but to each their own.
In size they fit somewhere between viruses and viroids (infections strands of RNA), and are rod-shaped, hence their name. And despite only being discovered recently, they are a lot of them. As the authors report: ‘Large scale searches identified 29,959 Obelisks … with examples from all seven continents and in diverse ecological niches.’
At this point, researchers don’t know if Obelisks are truly alive or not, what they evolved from, or if they are harmful or beneficial to their host organisms. In other words, stay tuned.
At the other end of the scale, a paper by two physicists from University College London suggest that dark energy and dark matter may not exist. To put this in perspective, under the currently most popular model of how the universe works – the lambda-CDM model (or, more simply, ΛCDM) – dark energy and dark matter make up nearly 95% of the universe. It’s like suggesting the theory of evolution through natural selection is fine except for the bit about natural selection … and maybe the bit about evolution.
Authors Jonathan Oppenheim and Andrea Russo ‘… consider a proposed alternative to quantum gravity, in which the spacetime metric is treated as classical, even while matter fields remain quantum.’ Making sense of this is way above my pay grade, but The Guardian’s science correspondent, Hannah Devlin, explains the theory this way: ‘(It) envisages the fabric of space-time as smooth and continuous (classical), but inherently wobbly. The rate at which time flows would randomly fluctuate … space would be haphazardly warped and time would diverge in different patches of the universe.’
If this sounds a bit like Doctor Who trying to explain time to Carey Mulligan’s character in the episode ‘Blink’, it may be because the universe is indeed ‘inherently wobbly’. It’s rather unsettling to think that the Weeping Angels might feel quite at home in Oppenheim and Russo’s universe.
Neither paper has been peer-reviewed at this point, but that hasn’t stopped them garnering media attention and commentary from other scientists. At the very least, ‘Obelisks’ and ‘wobbly spacetime’ have stirred the often lethargic currents of scientific orthodoxy; at best, they demonstrate that all scientific knowledge is provisional.
As FBI Special Agent Dana Scully says, ‘Mulder, the truth is out there.’
In part 4 of a thematic series of posts called Us, I said this about toolmaking:
‘It is with the application and development of tool usage that the first signs of a distinct ‘human’ culture are found in palaeoanthropology. Whereas chimps and some bird species, like humans, use tools made from plants to gather food or built shelter, humans are the first animals to make stone tools, improving on the original material through knapping.’[i]
Goffin’s cockatoo. Image courtesy of Creative commons.
I short-changed chimps, it seems. Not only do they use tools, they use tool sets: in other words they prepare different tools for different jobs.[ii]
More surprisingly, I now discover that I also short-changed birds. I was pleasantly surprised by the new(ish) information about chimps, but astounded by the news[iii] that at least one species of bird – Goffin’s cockatoo[iv] from Indonesia – also makes and uses different tools for different jobs.
I shouldn’t be astounded, of course. In a much earlier post I wrote about research providing evidence that corvids possessed a Theory of Mind. And as the article in The Conversation points out, an Australian bird – the palm cockatoo – is already known to regularly make drumsticks to beat against hollow trunks during courtship. I suppose it’s not a giant leap from all that to learning that at least one non-avian dinosaur could do with a tool box to keep its implements tidy and dust-free.
It seems that Goffin’s cockatoo actually manufactures three different tools – for wedging, cutting and spooning.
Again, as the article points out, this means the cockatoo’s cognitive skills can be compared directly with a chimps. Importantly, they have ‘… been confirmed as the third species that can not only use tools, but can carry toolsets in anticipation of needing them later on.’
The original research paper leading to the article in The Conversation can be found here in the journal Current Biology.
No, not another instalment in the Starship Troopers media franchise, but an exciting development in the war against the bugs that make us sick … and sometimes kill us.
Alexander Fleming’s discovery of penicillin in 1928 was a turning point in our struggle against bacteria-caused infection. Research carried out by Fleming, and subsequently by Cecil George Paine, Howard Florey and Ernst Chain, marked the start of the systematic production and use of antibiotics, at first in developed countries and later worldwide.
But after eighty years of use, antibacterial resistance is increasingly common. A 2014 report from the World Health Organisation states it is a threat ‘to global public health.’ The report found ‘high rates of resistance … in all WHO regions in common bacteria … ‘[i]
A 2016 review of antimicrobial resistance commissioned by the UK Prime Minister estimated that 700,000 people died each year from resistant infections, and that by 2050 ‘ … 10 million lives a year and a cumulative 100 trillion USD of economic output (could be) at risk … ’[ii]
So it’s kind of surprising that a paper published in February in eBioMedicine with the comparatively catchy title ‘A broad-spectrum synthetic antibiotic that does not evoke bacterial resistance’[iii] might garner some attention in the media. But there’s been hardly any attention at all, if any.
These few lines indicate why the paper may prove to be very important indeed in the future:
‘ … a promising compound, COE2-2hexyl, (exhibits) broad-spectrum antibacterial activity. (It) effectively-treated mice infected with bacteria derived from sepsis patients … including a CRE K. pneumoniae strain resistant to nearly all clinical antibiotics tested. Notably, (it) did not evoke drug resistance in several pathogens tested. (It) has specific effects on multiple membrane-associated functions … that may act together to abrogate bacterial cell viability and the evolution of drug-resistance.’
So not only did it treat bacteria (from sepsis patients) in mice, including a highly resistant strain, it did not evoke resistance and – importantly – might act against the evolution of drug resistance.
Hell, maybe COE2-2hexyl should feature in the next Starship Troopers movie. Sounds like it could take on any bug.
The remains of Homo floresiensis, discovered at Liang Bua on the Indonesian island of Flores in 2003, and of Homo naledi, discovered inside the Rising Star Cave in South Africa’s Cradle of Humankind, have played an important part in helping us understand the diversity and complexity of our hominin past.
Homo floresiensis. Photo courtesy of Creative Commons. Created by ATOR.
H. floresiensis, dubbed ‘The Hobbit’ by the media because of its diminutive size, with a brain capacity of around 380 cm3 and standing around a metre tall, was considered by many scientists to be a deformed or microcephalic H. sapiens. However, strong physical evidence such as humeral torsion[i] and a set of teeth unique among hominins[ii] has pretty well ended the debate about its status as a species in its own right. The main disagreement now, considering the size of its brain, is whether or not it should be included in the genus Homo.
And speaking of small brains …
H. naledi was half again as tall as H. floresiensis – about the same height as a large chimpanzee – and although its cranial capacity (between 460 cm3 and 610 cm3) was considerably bigger than the Hobbit’s, it was still well short of a modern human or any of our immediate cousins such as H. neanderthalensis.
Homo naledi. Photo courtesy of Creative Commons.
As I wrote in a previous post, however, brain size is not necessarily a reliable indicator of intelligence.[iii]H. floresiensis almost certainly made and used stone tools[iv], and recently the University of Witwatersrand’s Lee Berger announced that researchers had found evidence of fire being used by H. naledi[v]. This last was probably something of a given, since the remains of H. naledi were found in a chamber of the Rising Star Cave that could only be reached through a long, dark and twisting route that was difficult and dangerous to follow even with artificial light – without some kind of illumination it would have been virtually impossible. Still, this recent evidence adds weight to the case that this species was capable of making and using fire.
As friend and palaeoanthropologist Debbie Argue asks, however, when and how did H. naledi learn to make fire? Could they possibly have acquired the skill from a contemporary hominin, such as H. sapiens? Or was it the other way around? Or did both species learn the trick from a third hominin group?
We’ll probably never know the answer to this question, but it is fun thinking about, and – at the risk of stretching a metaphor almost to breaking point – throws another log on the fire of revaluating exactly what it means to be human.
[iii] (And in an even earlier post I write about evidence suggesting corvids, with comparatively lightweight brains (c. 20-25 grams, give-or-take), may have a Theory of Mind.)
One of the great palaeoanthropological bombshells of the last generation was the discovery of Homo floresiensis on the Indonesian island of Flores. For years scientists debated what ancestor this new and somewhat diminutive hominin – dubbed the ‘Hobbit’ by the media – had come from, or indeed if it should even be included in our genus.
Homo floresiensis reconstruction. Courtesy of Creative Commons. This image created by ATOR.
While now generally accepted as a member of our broader tribe, its origins are still fiercely argued, many insisting it’s nothing more than H. erectus that’s undergone insular dwarfism. But I think a 2017 paper written by Colin Groves, Debbie Argue, Michael Lee and William Jungers, convincingly demonstrates that H. floresiensis is not derived from H. erectus (or is a diseased example of H. sapiens), but rather from a much earlier hominim such as H. habilis or a sister species.[i]
A second paper, published in 2020[ii], backs up this hypothesis, and concludes with this statement:
‘ … something which on account of our inadequate current taxonomic framework we have to call “early Homo” differentiated in Africa, possibly as early as 2.8 (mya) … Subsequently, one or more members of this group reached the Mediterranean fringe and spread Out of Africa at 2.5 Ma. After successfully expanding over Asia, at least one of those hominins … gave rise to new species that reached the Caucasus by around 1.8 (mya), and thence Europe by ca. 0.9 (mya) … (the) eastward expansion (or occupation) in Asia of small-bodied and archaically-proportioned hominins continued, possibly in multiple waves; and, by ca. 0.8 (mya), representatives of this group had penetrated as far as insular southeast Asia, where H. floresiensis ultimately emerged … ’
Indeed, some scientists considered this possibility as early as 2005. A report about the brain of H. floresiensis published in Science in that year[iii] concludes with these lines: ‘Although it is possible that H. floresiensis represented an endemic island dwarf that, over time, became subject to unusual allometric constraints, an alternative hypothesis is that H. erectus and H. floresiensis may have shared a common ancestor that was an unknown small-bodied and small-brained hominin.’
Homo habilis. Courtesy of Creative Commons. Photographer unknown.
I think an increasing weight of evidence strongly suggests that the first major exodus of our genus from Africa was carried out by H. habilis or one or more of her sisters. Furthermore, I think it’s possible that these closely related species then gave rise to H. erectus, H. pekinensis, H. luzonensis[iv] and H. floresiensis in Eurasia, while those remaining in Africa gave rise to H. ergaster. This does not preclude the possibility, or perhaps probability, of any or all of these species crossbreeding if they ran across each other.
But what of H. sapiens, our own species? As with H. ergaster and H. erectus, the evidence here is convoluted, confusing and often contradictory.
Mongrel
For those, like Colin Groves, who think H. ergaster is a species in its own right, the line of descent works something like the following.
Homo heidelbergensis. Courtesy of Creative Commons. This image created by ATOR.
About 600,000 years ago, H. ergaster, either directly or through an intermediary species called H. rhodesiensis, gave rise to H. heidelbergensis. This species was our size physically, and his brain capacity was well inside the standards of Anatomically Modern Humans (AMH). Following the great tradition of hominin migration, something that seems as ingrained in our genus as bipedalism, some members of this new species moved to Europe[v]. About 400,000 years ago, they gave rise to H. neanderthalensis. In a case of ‘well, we’ll show you’, those who stayed behind in Africa gave rise to H. sapiens at least 300,000 years ago, and possibly as long as 350,000 years ago.[vi]
I can’t stress this enough. Homo sapiens are Africans. It is where our archaic ancestors and AMH first appear[vii]. (Let me also stress that this story, as complicated as it gets from now on, does not resurrect the Multiregional Model for our evolution, where H. erectus gave rise to H. sapiens across its whole range at the same time, from Africa to Asia. This is an old theory, now largely discredited by the extensive fossil and DNA evidence that our species first evolved in Africa.[viii])
What happened next has been slowly and painstakingly uncovered by palaeoanthropologists doing field work throughout Africa and Eurasia, and by the outstanding work performed at the Max Planck Institute’s Department of Evolutionary Genetics, headed up by Svante Pääbo, into hominin DNA.[ix]
What the DNA evidence strongly suggests is that H. sapiens successfully left Africa between 70,000 and 100,000 years ago. (Although this wasn’t the first migration into Eurasia by our species. It is usually held that previous attempts left no trace in the DNA of AMH outside of Africa, but see these earlier posts, here and here.)
Female Homo neanderthalensis. Courtesy of PLOS ONE.
Members of the most recent migration interbred with H. neanderthalensis, probably in what is now the Middle East, and later with the Denisovans, another possible descendant of H. heidelbergensis, deeper in Eurasia[x]. To this day, the average ex-African H. sapiens carries between 1%-2% of the Neanderthal genome; but it is not the same one or two percent: we overlap. Overall, we carry up to 40% of the Neanderthal genome in our own genes. But the story gets more complex still: the genome of people from Oceania, such as Papuan New Guineans and Australian Aborigines, can have between 5-6% Denisovan DNA[xi]; indeed, recent research suggests that Ayta Magbukon Negritos in the Philippines have Denisovan ancestry 30-40% higher than either of these two groups.
The Natural History Museum of London’s Professor Chris Stringer says, ‘It is now clear there was a lot more interbreeding between ancient species, including early Homo sapiens and others, and that there was a lot more movement of populations both in the distant past – and relatively recently.’[xii]
Homo sapiens (Oase 2) reconstructed from bones 37,000-42,000 years old discovered in the cave of Peştera cu Oase in Romania. Around 7.3% of his DNA is from H. neanderthalensis, from an ancestor 4-6 generations back. Courtesy of Creative Commons. Photo: Daniela Hitzemann.
Talking about recent research, in June last year Chinese scientists announced that a cranium first discovered in China almost a century ago, is a new species of Homo with a brain easily the equal of any AMH in size and carried inside a skull more massive than ours. Those making the announcement have named the new species H. longi (‘Dragon man’, and just as Denisovans are sometimes described as a sister species to Neanderthal, so H. longi is being claimed as a sister species to H. sapiens[xiii]).
As Lee Berger, from the University of Witwatersrand and the discoverer of Australopithecus sediba and H. naledi, has suggested, perhaps the different paths of human evolution are not best thought of as branches spreading from a single tree trunk, or even a messy, many-twigged bush, but rather a braided stream[xiv] with tributaries constantly running across each other before separating, rejoining and separating once more.
The Waimakariri River in New Zealand is braided along almost its entire length. A good metaphor for hominin interbreeding? Courtesy of Creative Commons. Photo: Greg O’Beirne.
We, Anatomically Modern Humans, are the result of all this evolution. We are nothing more than a mongrel species.
And this from the Australian Museum: ‘Most scientists that accept H. floresiensis as a legitimate species now think its ancestor may have come from an early African dispersal by a primitive Homo species similar in appearance to H. habilis or the Dmanisi hominins. This means that it shared a common ancestor with Asian H. erectus but was not descended from it. Cladistic analysis supports the lack of a close relationship with H. erectus.’
[v] The first H. heidelbergensis fossils were found near Heidelberg in 1907.
[vi] Although this paper suggests the split between our two species might be found much further back … up to 800,000 kya or more!
[vii]Recent research from scientists at Australia’s Garvan Institute of Medical Research reveals that southern Africa is home to the oldest evidence for AMH: ‘… to contemporary populations that represent the earliest branch of human genetic phylogeny.’ The date they arrive at is 200,000 years ago.
As well, a report in the February issue of Science describes how thousands of genome sequences were collected from modern and ancient humans to create a family tree. In the words of the report’s first author, Anthony Wilder Wohns, ‘ … we definitely see overwhelming evidence of the Out-of-Africa event … ‘
[viii] See Stringer, C. & Andrews, P. The Complete World of Human Evolution. London, 2011. P 140 ff for a discussion of the two main theories for the evolution of Home sapiens: ‘Multiregional’ and ‘Out of Africa’.
[ix] And now, besides DNA, they are using protein analysis to identify ancient hominins, most recently the first Denisovan found outside of the Denisova Cave in Siberia … on the Tibetan Plateau of all places! See https://www.nature.com/articles/s41586-019-1139-x, 16 May 2019.
[x]Very recently, H. sapiens remains were discovered in the Grotte Mandrin rock shelter in the Rhône Valley in France that date back 54,000 years ago, pushing back our species arrival in Europe by at leat 10,000 years from previous estimates.
[xi] Please watch this fascinating talk Svante Pääbo gave at the University of California in 2018 after receiving the Nierenberg Award for Science in the Public Interest. It goes into all of this in much more detail. As Pääbo points out in the talk, the DNA evidence indicates humans ‘have always mixed’.
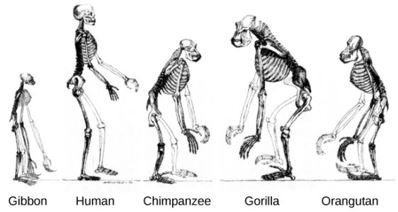
Humans walk upright, gorillas and chimpanzees walk on all fours, resting their weight on their knuckles, and orangutans can do just about anything – they hang and swing by their arms from branches, sometimes with the help of their oddly-shaped feet, and on the ground they can walk either upright or on all fours. The structure of the postcranial skeleton in all four animals is very different and reflects these locomotor patterns. Non-human great apes have short legs and long arms, whereas we have very long legs. With the gorilla and chimpanzee it is the shortness of the legs that differs from humans, the arms being much more similar in length compared to the torso; only the orangutan has enormously lengthened arms. When other great apes stand upright, their legs are straight from hip to ground, whereas humans are ‘knock-kneed’, as the thighs slope inward from the hip to the knee. The pelvis is very different in appearance: in humans the hip bone (ilium) is low and very broad, but in great apes it is high and fairly narrow. In humans the great toe is long and stout and aligned with the other toes, but in great apes it is divergent from the other toes (less in the gorilla), and in the orangutan it is very short.
Courtesy of Creative Commons. Artist unknown.
In most great apes, the spinal column is more or less straight, but in humans the spine is curved into a double-S: the cervical (neck) vertebrae curve forward, the thoracic (chest) vertebrae curve backward, the lumbar vertebrae (those in the small of the back) curve forward again, the sacral vertebrae (which are fused together, and form the back wall of the pelvis) curve back again, and the coccyx (the partially fused vertebrae which are the tiny remnant of the tail) curves forward once more. The ribs (which are very variable in number, but average 12 in humans and orangutans, and 13 in chimpanzees and gorillas) together form the thorax; in humans the thorax is barrel-shaped (narrow at the top, broad in the middle, narrower again at the bottom), whereas in great apes it is funnel-shaped (narrow at the top, and broadening towards the bottom).
All of these differences between humans and the other great apes are developments stemming from bipedalism. So why did humans adopt bipedalism? Well, walk with me and we’ll take a brief look at the major theories.
Doing a runner
There seems to be a growing consensus among many scientists that our ancestors evolved bipedalism for several reasons rather than one overriding factor. What many of the competing theories do agree on, however, is that rainforest giving way to savannah because of climate change around 7.0 – 5.0 mya was a strong influence. Grassland with only scattered trees and no closed canopy meant tree-climbing primates had much more open territory to cover. Walking on two legs freed hands to carry infants, food or tools, including weapons. Walking on two legs made us taller, meaning we could locate food, potential predators and safe havens from further away; it also made it easier to pick low-hanging ripe fruit from trees. Walking reduced the amount of body surface area we exposed to the sun while in the open.
Early morning on the savannah. The change in the landscape from rainforest to savannah between 7 mya to 5 mya probably helped kickstart bipedalism in hominins. Photo: Simon Brown.
Of course, in some circumstances some of these ‘advantages’ could become disadvantages. For example, although bipedalism meant we could locate a predator from further away, it also meant if it was looking in the right direction, a predator could see us from further away as well (and our chief predators – leopards, hyenas and lions – all have good eyesight, not to mention excellent hearing and sense of smell). On the other hand, when our ancestors became active hunters, our extra height gave us an advantage over prey animals, many of whom rely on their sense of smell rather than their eyesight.
Our genus has evolved to become a natural endurance runner, and through that a natural persistence hunter. Courtesy of Creative Commons. Photographer unknown.
More recently, one of the major arguments for the successful adaptation of bipedalism was that it is a much more energy efficient method of locomotion[i]. Whatever the arguments for or against all these hypotheses regarding the origins of walking, when it came to running there is no denying our bodies evolved to make us one of nature’s supreme endurance runners[ii]. This seems to have happened about two million years ago and was a real game-changer when it came to predating: our ancestors evolved into persistence hunters, able to wear down much larger animals such as kudu and oryx[iii]. Basically, humans ran their prey into the ground, and much of our body shape is particularly adapted to long-distance running.
In other words, the characteristics that make us superb walkers and runners are the characteristics that most set us apart from other great apes. As Chris Stringer and Peter Andrews write in The Complete World of Human Evolution, ‘at present … (bipedalism) is taken as the earliest adaptation by which we can recognise human ancestors in the fossil record.’[iv]
The odd-sock drawer
Now it’s time to deal with one of the most controversial species in the human lineage – Homo ergaster. This species was described by Colin Groves and Vratislav Mazák in 1975[v]. Since then, palaeoanthropologists are divided on whether H. ergaster is a distinct species, or a subspecies belonging to H. erectus, palaeoanthropology’s pin-up boy and all-purpose species.
Once they learned to walk, our ancestors just kept on walking. In fact, they walked right out of Africa, into the Middle East, then east into Asia and Sahul, north to Europe, and eventually across the Bering Strait and into the Americas. On the way they continued evolving into new species that seemed to interbreed with each other at every opportunity, creating yet more new species, and eventually discovering agriculture, television and the internet. And interestingly, it’s the use of technology that provides us one piece of evidence that H. ergaster and H. erectus were two different species.
But first, let’s talk more about bones, specifically those belonging to the original H. erectus, parts of which were first discovered 1891 by Eugène Dubois, a Dutch doctor working for the army in Java. In fact, he went to Java with the objective of discovering evidence supporting the theory that H. sapiens evolved in Asia, an idea most determinedly supported by German naturalist Ernst Haeckel. Haeckel had hypothesised that our species’ progenitor, which he names Pithecanthropus alalus, had evolved on Lemuria, a mythical continent that subsequently sunk beneath the Indian Ocean (thereby conveniently leaving no fossils behind to prove – or for that matter, disprove – his theory).
Eugène Dubois, discover of Homo erectus. Courtesy of Creative Commons. Photographer unknown.
Although Dubois had discovered ancient hominin fossils, he found little or no support among scientists in Europe that they amounted to anything significant. It wasn’t until Sinanthropus pekinensis was discovered in China over a quarter-century later that enthusiasm for Dubois’s discovery really picked up. In the early 1950s, Ernst Mayer reclassified both P. alalus and S. pekinensis as H. erectus[vi]. Since then, hominin fossils with roughly the same estimated brain size as H. erectus and aged between 2 million years old to just over 100,000 years old have been thrown in with H. erectus like differently coloured socks thrown into an odd-sock draw. It has become the species to have when you want to cover all of Africa and Eurasia and two million years of history.
In the early 1970s, for example, Richard Leakey and Alan Walker described two partial skulls found in Kenya as belonging to an African offshoot of H. erectus based on the fact that their calculated brain capacities (848 cc and 803 cc) were not dramatically smaller than that of some H. erectus skulls (around 950 cc), which is like arguing that since the Volvo S60 and the Volkswagen Passat have similar interior space, they’re both examples of a Toyota Camry.
However, in 1975, Colin Groves and Czech colleague Vratislav Mazák, after a comprehensive metric analysis of fossils from Koobi Fora, discovered they had uncovered a new species they names H. ergaster. Their argument was that there was no African version of H. erectus; further, Colin Groves believed that H. ergaster evolved in Africa and then migrated into Eurasia, eventually giving rise to H. erectus.[vii] The earliest dates for the new species goes back 1.9 million years[viii], as opposed to 1.6 million years (or 1.8 according to some estimates) for H. erectus, making H. ergaster the first truly human-looking hominin to stride the planet – tall, thin, decidedly bipedal, with a flatter face than its ancestors and an active hunter, fire-user and tool-maker.
KNMER 3733, possible cranium of a female Homo ergaster. Photo: Simon Brown.
Now, nearly fifty years after the initial paper by Groves and Mazák, a fierce debate still continues between those who think the two hominins are separate if linked species, or just subspecies. In common parlance, it’s a debate between splitters and lumpers.[ix]
But besides the obvious difference in the skull shapes of H. ergaster and H. erectus, another line of evidence convinces me that Colin was right in his opinion that we are talking about two species. This evidence involves tool making.
Out with the old, in with the new
Until the appearance of H. sapiens and H. neanderthalensis, stone age technology is divided into two broad and overlapping stages: Oldowan and Acheulean (sometimes called Modes 1 and 2). Oldowan technology was first discovered in the 1930s by Louis Leakey at the Olduvai Gorge in Tanzania. The oldest examples have been found at Gona in Ethiopia, and date back about 2.5 million years[x]. The technology seems to have spread very quickly, and recent discoveries have found stone tools in Jordan dated at 2.5 mya and China at 2.1 mya[xi]. This technology, the use of very simple flakes and rocks, had been developed before the appearance of H. habilis, possibly by Australopithecus garhi. Acheulean technology which started about 1.76 mya, is closely associated with the appearance of H. ergaster and involves more refined knapping and the development of specialised tools such as hand axes.
This doesn’t imply that Oldowan technology suddenly evaporated, and every hominin adopted the new style of knapping chert. In some places, Oldowan and Acheulean stone tools are found at the same site from the same period, suggesting that while H. ergaster or one of its descendants employed the improved technology, one of our cousins continued using the older method. But it’s clear Acheulean technology obviously conferred a significant advantage over the old style. It didn’t take long for it to spread beyond Africa, either because H. ergaster itself started spreading beyond Africa, or because it spread by ‘word-of-mouth’: neighbouring hominis picked up on the new fashion of making tools and copied it. Acheulean tools appear in what is now India, for example, by 1.5 mya, and in Europe by about 900 kya.
Acheulean hand axes. Compare the careful knapping done here to the more primitive Oldowan tools illustrated in the previous post. Courtesy of Creative Commons. Photographer unknown.
However, Acheulean technology did not seem to reach Java, where our friend H. erectus resided.
Which presents lumpers with a problem. If H. ergaster is indeed nothing more than a subspecies of H. erectus, then fossil evidence suggests this single species arose in Africa before spreading throughout Eurasia. Yet if this is also the species that developed Acheulean technology soon after evolving, why didn’t the technology travel with them to the far east?
On the other hand, if we are talking about two species, then it’s quite possible for Acheulean technology to be developed by H. ergaster in Africa, spread slowly throughout Eurasia, but never quite reach the home of H. erectus in Java.
If this was in fact the case, it raises a more important question: even if we accept H. ergaster is a separate and earlier species than H. erectus. Does it necessarily follow that H. ergaster gave rise to H. erectus? What if the two species are cousins rather than mother and daughter?
This is something we’ll discuss in the next, and final, post of ‘Us’.
‘(He) was the first to suggest that the genus Pithecanthropus should be subsumed into Homo, and in the same paper he proposed that fossils recovered from what was then called Choukoutien (now called Zhoukoudian), which were initially assigned to Sinanthropus pekinensis,26 should also be transferred to H. erectus.’
‘A growing number of scientists have redefined the species Homo erectus so that it now contains only east Asian fossils. Many of the older African fossils formerly known as Homo erectus have now been placed into a separate species, Homo ergaster and this species is considered to be ancestral to Homo erectus. The redefined Homo erectus is now generally believed to be a side branch on our family tree whereas Homo ergaster is now viewed as one of our direct ancestors. ‘
[viii] Oldest fossil dates according to the Australian Museum for H. ergasterhere and for H. erectushere. Recent work reported in the journal Science may push the dates even further back, between 1.95-2.04 mya (although in this paper the discussed specimen is describe as preserving ‘characters that align it morphologically with H. erectus sensu lato (including Homo ergaster)’. Go figure.
[ix] For a fuller description of the often heated debate about what makes a species, see here.
[x] Stringer, C. & Andrews, P. The Complete World of Human Evolution. London, 2011. P 208.
A new kind of stone age technology – Lomekwian – has been suggested after the recent discovery of stone tools at Lomekwi that predates Oldowan by more than 700,000 years. See the previous post for more details.
Forgive the pun, but for decades it seemed a no-brainer that the chief qualification to be considered human was the size of your brain. Obviously, it had to be a of a certain respectable capacity, never quite defined, but a degree or two larger than a chimpanzee’s organ was a good start. There was some embarrassment when it was determined that the average brain capacity of Homo neanderthalensis was larger than our own[i], but that misgiving aside it was assumed that if not a directly comparative intelligence was a prerequisite, then certainly something within shooting distance.
One mistaken assumption here is between brain size and intelligence, something made very clear in recent years by the discovery of the stone tool-making H. floresiensis (with a brain the size of a chimpanzee). Recent work done on corvids, for example, suggests that ravens and crows possess a Theory of Mind[ii] – the capacity to imagine that another crow might have its own thoughts – which in turn suggested a reasonably developed sense of self-awareness, an emergent property traditionally associated with intelligence[iii].
Homo heidelbergensis had a brain around 1200 cc, well within the range of H. sapiens. Photo: Simon Brown.
Another mistaken assumption is that our larger brain size is extraordinary among our cousins, but average brain size has not increased dramatically in total capacity since H. heidelbergensis, a species that first saw light of day 600,000 years ago.[iv]
Indeed, Homo species sit comfortably on the line that matches a generic primate’s brain size to its body size. In other words, if you’re a primate, the bigger you are the bigger your brain gets. (This isn’t peculiar to primates, of course, and applies to many mammalian groups, eg rodents, elephants and aardvarks, but primates do have larger brains than mammals of similar body mass).[v]
Interestingly, there are three exceptions to this general rule, all three of which are closer to us genetically than any other primates: the orangutan, the chimpanzee and the gorilla. The orangutan falls just below the curve, the chimpanzee falls a little further, and the gorilla furthest of all. Extensive studies with chimpanzees and gorillas, however, show that both species are intelligent and self-aware enough to have developed a Theory of Mind.
Demonstrably, brain size is not irretrievably married to a set physical size, just as brain size is not irretrievably married to a set level of intelligence.
I know that you know that I know …
It does seem self-awareness, or sentience, is an emergent property of intelligence.[vi] In other words, as an animal increases in intelligence, at some point it will become aware of its own existence. This is more than simply being able to experience pleasure or pain, but the ability to experience life subjectively.
Objects found with the remains of H. floresiensis strongly suggests they made stone-age level weapons and tools.[vii] Obviously, such complex toolmaking suggests an active intelligence capable of learning new skills and – as importantly – passing those skills on to the next generation. This in turn suggests H. floresiensis possessed a language; if not a spoken language such as ours, with a huge vocabulary and complex rules of grammar, then at least some way to transmit a limited amount of information effectively and efficiently.
Evidence also exists that H. floresiensis hunted and scavenged animals such as the dwarf stegodon, a kind of elephant. To be clear, a dwarf elephant could still grow to more than two metres in height. For something the size of H. floresiensis to hunt stegodon strongly suggests they hunted in groups, which in turn strongly suggests their language was something more than a series of grunts.
With H. naledi, we are on somewhat less firm ground. Although they were larger-brained hominins than H. floresiensis, the remains of at least 15 individuals from the Rising Star Cave in South Africa were discovered without any tools or evidence of tool making. However, the species possessed a hand not dissimilar to our own, and would probably have been capable of tool-making. It is hard to imagine a hominin species living in Africa in this period, between 236,000 and 335,000 years ago, and not picking up the skill from one of the other hominin species occupying southern Africa at the same time (including, quite possibly, our own).
The Rising Star Cave in South Africa, where the fossils of several Homo naledi were discovered in the Dinaledi Chamber. H. naledi almost certainly would have needed fire – and a great deal of determination – to find their way to the chamber from its entrance. Courtesy of Creative Commons.
Furthermore, palaeoanthropologist Lee Berger, who led the expedition to recover the H. naledi remains from the Rising Star Cave, believes bodies were intentionally and repeatedly deposited there. This implies two things: first, ritual behaviour on the part of the species, and second, that they were capable of making fire, since the chamber the bones were discovered in is at the end of a long, dark, dangerous and narrow route.[viii]
I may not know much about art, but …
In his influential work on human development, The Ascent of Man, Jacob Bronowski wrote, ‘Man is not the most majestic of creatures. Long before the mammals even, the dinosaurs were far more splendid. But he has what no other animal possesses, a jig-saw of faculties which alone, over three thousand million years of life, make him creative. Every animal leaves traces of what it was; man alone leaves traces of what he created.’[ix]
Whether or not Bronowski used the term man to mean, specifically, H. sapiens, or more broadly to mean humans in general, we know that our cousins left behind more than traces of what they created. We have hundreds of stone tools, the tailings and debris of stone-tool manufacturing, and even examples of art.
I would suggest this equates to culture.
But what if there are no physical signs of culture, does it mean culture does not exist? It is often fallacious to argue that absence of evidence is not evidence of absence, but in cultural endeavours such as language or dance, there can be no evidence before the invention of writing and art.
Vervet on the lookout for predators. Photo: Simon Brown.
Simple language can be identified in many primates. Vervet monkeys, for example, have distinct calls for each of their four main predators: pythons, baboons, leopards and eagles. But we will probably never know which human species was the first to communicate with what we would describe as a complex language, one capable of conveying abstract thought. Vervet monkeys may be able to tell their fellows that a leopard is approaching, but they cannot say the leopard is hiding behind that bush or over that hill, let alone discuss the rights and wrongs of predation.
We see culture operating among our more social hominid cousins, the chimps and gorillas. Long-term field studies suggest, for example, that cultural variation exists among different chimpanzee groups, including differences in grooming, courtship and tool usage.[x] It is the ‘combined repertoire’ of chimp behaviours that is significant, demonstrating a range of cultural behaviours, a diversity that once was attributed only to our own species.
It is with the application and development of tool usage that the first signs of a distinct ‘human’ culture are found in palaeoanthropology. Whereas chimps and some bird species, like humans, use tools made from plants to gather food or built shelter, humans are the first animals to make stone tools, improving on the original material through knapping. Later, humans combined stone with other material, such as wooden handles, to improve their effectiveness; in other words using tools to make better tools. Indeed, the making of stone tools was once considered the boundary marker between members of Homo and earlier genera. Since then, the boundary for stone-tool making has been pushed well beyond those species traditionally grouped under our own genus.
Oldowan stone age tools. Courtesy of Wikimedia Commons. Photographer: Didier Descouens.
The oldest crafted stone tools found so far are from Lomekwi in Kenya, dating back 3.3 mya[xi]. First discovered in 2011, they were probably made by a species belonging to either the Australopithecus or the Kenyanthropus genera. The tools were found in an area where Kenyanthropus platyops fossils had been found earlier.
But we have to wait more than 700,000 years before there is clear evidence of stone-tool making on a large scale, something we’ll cover in detail in a later post.
Eventually some hominins were not simply making stone tools: ‘The people who made the hand axes clearly had a specific shape in mind, and often went far beyond a purely utilitarian form in the care with which they produced them.’[xii] This is an example of humans crafting tools for aesthetic appeal, not just knapping to produce a sharp edge or a convenient grip.
Earliest example of art was created by Homo erectus 500,000 years ago. Courtesy of Creative Commons. Photographer unknown.
It is with H. erectus we find the first real example of an attempt at making what we would now call ‘art’. In 2014, scientists from Netherland’s Leiden University announced the discovery of a sea shell that had been engraved with a zigzag pattern 500,000 years ago, something identified by ANU scientist Dr Stephen Munro (who did his PhD under Colin Groves!). The shell was originally collected with others at the end of the 19th century by Eugène Dubois – the discoverer of H. erectus in Java – but had not been closely examined since the 1930s. The scientists demonstrated that not only was the engraving not the result of natural forces, but that the pattern was made by ‘a strong and skillful tool-maker’[xiii]. The new date pushed back the first evidence for art by 400,000 years.
With language we’re on much shakier ground. Research suggests the physiological requirements for language exist in at least some monkeys. The stumbling block seems to arise in the way the brain is wired[xiv].
Nonetheless, as noted above with vervet monkeys, a language with a basic vocabulary exists among many primate species. It has even been shown that different species of monkey may understand some of each other’s vocabulary[xv]. Some species have even developed a basic grammar[xvi].
Extensive work has been done on language among the great apes, both in the wild and under controlled conditions. For example, the remarkable success scientists have had teaching American Sign Language to Washoe, a chimpanzee, and Koko, a lowland gorilla, demonstrate their capacity to learn quite complex vocabulary, often using it to express emotions such as sadness.
Koko, a western lowland gorilla, with her pet cat All Ball. Courtesy of Creative Commons. Photographer unknown.
But even the most optimistic view of these experiments shows that non-human great apes never demonstrate a level of intelligence found in a three-year old human child. No chimpanzee or gorilla, for example, has ever used their acquired vocabulary to ask a question.[xvii]
There is genetic evidence to suggest that the development of the capacity for language accelerated in humans after we split from the chimpanzees some seven to eight million years ago[xviii], but precisely when humans started speaking in a way that we would describe as ‘human’ is unknown; it may never be known. As with so many things in evolution, the development of a complex language capable of expressing abstract thoughts almost certainly occurred along a spectrum.
Between them, language and craft handed humans a huge advantage in the evolutionary stakes. Making stone tools, for example, minimised our weaknesses, knives and hammers allowing us to make up for a lack of sharp claws and fangs. Later, bows and throwing spears made up for our lack of speed in the chase.
Language allowed us to magnify our strengths, especially the ability to learn new things and pass that learning on to succeeding generations.
Language, and culture generally, seems to be something we share with other members of our genus, and indeed, as they are presently classified, earlier genera.
In the next post we’ll talk about bipedalism and one of the most controversial of hominin species – H. ergaster.
[i] Specifically, larger on average than the modern human brain, although the brains of archaic H. sapiens were in fact comparable to H. neanderthalensis. The following excerpt is from here.
‘To measure fossil brain volume, anthropologists have traditionally filled skulls with beads or seeds, and dumped the contents into a graduated cylinder (a precise measuring cup). They’ve also submerged molds of skulls into water, measuring the volume displaced. Today CT (computed tomography) scanning methods offer more accurate (and less-messy) measurements, but much of the data in textbooks and other references was collected the old fashioned way.
‘Based on these values, we can confidently say fossil Neanderthals and modern humans from the same time period had similar brain sizes. Twenty-three Neanderthal skulls, dating between 40,000 and 130,000 years ago, had endocranial volumes between 1172 to 1740 cm3. A sample of 60 Stone Age Homo sapiens ranged from 1090 to 1775 cm3.’
[xvii] Some scientists argue that Koko’s language skills were a result of ‘operant conditioning’, whereas others state she was indeed capable of simple questions. See Wikipedia entry here for more information and references.
Most of us did some biology at school, and most of us came out with the idea that species are groups of populations that cannot interbreed. When we’re reminded of mules, which are the offspring of horses and donkeys, we think ‘Ah, but they are sterile, aren’t they?’ Almost invariably, although there have been a few cases of fertile mules, but when cattle and bison interbreed, while the male offspring are sterile, the female offspring are fertile. All the big cats can also mate with each other, producing hybrids (where the female is fertile), and in the case of the leopon, the hybrid between a leopard and a lion, even the male might be fertile.
A tigon: hybrid from tiger father and lion mother. Because it’s a male, it would be infertile. Courtesy of Creative Commons. Photographer unknown.
Now that we can trace the ancestries not only of individual people, but whole populations and whole species, through DNA, it turns out that there has been a whole lot of successful – that is, fertile – interspecies breeding in the past. And it sometimes turns out that different species even today may interbreed with their neighbours on the quiet. For example, the primatologist Kate Detwiler discovered that two species of small monkeys in Tanzania’s Gombe National Park, the red-tailed monkey (Cercopithecus ascanius) and the blue monkey (Cercopithecus mitis), are found living in separate troops in some of the forested valleys, but in other valleys interbreed – in fact, in one or two valleys the monkey population consists entirely of hybrids.[i]
The idea that different species don’t interbreed is simply not true. They may not do so usually – but that is another thing entirely. We cannot use non-interbreeding as a criterion for species.
How then, can we define species?
George Gaylord Simpson. Courtesy of Creative Commons. Photographer unknown.
For over 150 years now, the basic guiding principle of biology has been evolution – so the question we should be asking is what is the evolutionary status of species? The palaeontologist George Gaylord Simpson (1902-1984) suggested in 1961 that the essence of species is that they are evolutionary lineages. He got little reaction at the time because his colleagues largely were hung up on the non-interbreeding criterion, but from the late 1990s his insight has been more and more appreciated. The best way to recognise an evolutionary lineage is, quite simply, that it differs from other evolutionary lineages. Horses and donkeys differ consistently and therefore represent two separate evolutionary lineages, and are therefore two different species. Similarly, blue monkeys and red-tailed monkeys differ consistently and therefore constitute separate evolutionary lineages, and again represent two different species.
Stumptailed macaque. Courtesy of Creative Commons. Photographer unknown.
If there are whole populations which consist of hybrids between two species, then what? Sometimes hybrid populations remain isolated for a good length of time and become homogeneous – and a new species is born. At least one species of monkey, the stumptailed macaque (Macaca arctoides) of mainland Southeast Asia, is thought to have arisen about 1 million years ago from a hybrid population between two other species.[ii]
For a more detailed discussion about the arguments about how to define species, especially the contest between the Biological Species Concept and the Phylogenetic Species Concept, go here.
So … generally speaking, what are genera?
So what about genera, families and other taxa?
While the taxa at both ends of the ranking are pretty straight forward – ‘species’ is eminently useful, and ‘domain’ and ‘kingdom’ are irresistibly sensible – all the ranks in between can get awfully confusing. And they are actually rather arbitrary. When, for example, do we know that a group of organisms constitute a genus rather than a family?[iii]
One simple solution would be to organise those in-between ranks chronologically. In other words, the order Primates would include all those monkey, ape and human-like species which existed from the Palaeocene epoch, and the family Bovidae would include all those antelope, buffalo and cattle, and sheep and goat species which existed from the early Miocene epoch.
This is an idea first forcefully proposed by German biologist Willi Hennig (1913-1976), considered the founder of cladistics – or ‘phylogenetic systematics’ if your thesaurus is turned on.
Willi Hennig. Courtesy of Creative Commons. Photographer unknown.
In 1966, Hennig proposed linking the taxonomic rank of a clade to its time of origin. He argued that if taxa are to mean anything they must represent monophyla – that everything in that group must be descended from a common ancestor. He also argued that taxa had to be characterised chronologically.
Hennig was an entomologist and realised while many genera of insects separated from one another tens of millions of years ago, the genera of mammals and the genera of birds separated more recently.
The idea was taken up by American scientist Morris Goodman (1925-2010), one of the founders of molecular genetics. He set about constructing a consistent scheme for the group of mammals about which he was most familiar – the primates. In 1997, he suggested that a reasonable time depth for a primate genus would be seven million years, partly because this would do the least violence to the presently accepted system of determining genera.
Colin surveyed many of the mammalian genera that taxonomists had recognised and found that most had separated from each other less than seven million years ago. Subsequently, he proposed that five million years was a more appropriate time depth for mammalian genera: the Miocene-Pliocene boundary.
Furthermore, Colin suggested that the taxonomic rank of ‘family’ had a time depth of 24 million years, separate families splitting around the time of the Oligocene-Miocene boundary. Going up one more ranking, the different ‘orders’ separated around the time of the Cretaceous-Tertiary boundary (the famous K-T boundary that marks the arrival of the asteroid that wiped out non-avian dinosaurs[v]).
Colin Groves. Photo: Simon Brown.
One of the consequences of Goodman’s proposals for palaeoanthropology is that most if not all members of the human lineage would belong to a single genus. Indeed, using his original suggested time depth of seven million years, Goodman even included chimpanzees into Homo. Overall, the later modifications devised by Colin play less havoc with the established order, but they would still require that most human fossils be placed in the same genus as ourselves.
Arguments about when hominins evolved into a genus that can be described as wholly human traditionally revolved around the relative importance of different physical characteristics: brain size, dentition, general morphology (body size, especially the extent of sexual dimorphism), and primary form of locomotion.
Other more controversial factors sometimes taken into consideration include tool-making, art and other signs of culture, and evidence of community living.
For example, some experts such as Ian Tattersall, curator emeritus with New York’s American Museum of Natural History, argue that the cranium of Homo floresiensis (the Hobbit, see here, here and here) is too archaic for it to be included in our genus.
This leads us to our second, and more controversial opinion: following Colin’s plan our genus would include not only H. floresiensis but even older and more archaically featured species traditionally belonging to other genera, such as the Australopithecines, for example, which include the Taung Child and Mrs Ples.
Colin argued that the Miocene-Pliocene boundary more or less corresponds to the onset of the only characteristic definitely belonging solely to our genus and to no other genera among the great apes – bipedalism. By bipedalism we mean that the main form of locomotion is walking or running on two legs, with the big toe aligned with the other toes in the foot.[vi]
Accepting this argument has two major implications and several minor ones for palaeoanthropology. First, and least controversially, brain size is not by itself a qualification for membership of the human genus. Specifically, a small brain does not exclude membership.
Homo naledi. Courtesy of Creative Commons. Photographer unknown.
The discovery of H. floresiensis and H. naledi in the 21st century, with an average brain size of around 420 cm3 (about the size of a modern chimpanzee) and 500 cm3 respectively, clearly demonstrates that many humans were small brained compared to H. sapiens but possibly still capable of sophisticated tool-making and ritual behaviour.
Secondly, accepting a time criterion in determining what species do and do not belong to the genus Homo means that strictly morphological traits are no longer intrinsic in determining human status.
In the next post, we’ll look in more detail at brain size, culture and bipedalism as criteria for determining whether or not a species is human.
[iii] Sigwart, J., Sutton, M. D., & Bennett, K. (2017). ‘How big is a genus? Towards a nomothetic systematics’. Zoological Journal of the Linnean Society. https://doi.org/10.1093/zoolinnean/zlx059
[iv] Groves, Colin. ‘Time and taxonomy’. Ludus Vitalis. Vol IX. No 15. 2001.
& Groves, Colin. ‘Speciation in hominin evolution’. African Genesis: Perspectives on Hominin Evolution. Ed Reynolds, Sally & Gallagher, Andrew. Cambridge University Press. 2012.
& Groves, Colin. ‘Current taxonomy and diversity of crown ruminants above the species level’. Zitteliana B32, International Conference on Ruminant Phylogenetics, ed. Prof. Dr G Worheide, Bavarian State Collection for Paleontology and Geology, Munich.
[v] Now also sometimes referred to as the Cretaceous-Palaeogene (K-Pg) boundary.
‘Burdalone’ is an old Scottish word meaning the last bird in the nest, the one left when all the other chicks have flown or all the other chicks have died. It’s a sad and lonely word, and perfectly describes Homo sapiens.
When one of the first members of our own species studied the world around her, most of what she saw would be familiar to us today, whether from personal experience or from watching nature documentaries about Africa. Extensive grasslands dotted with acacias, watering holes and narrow rivers with crumbling banks, herds of large grazing animals such as wildebeest and zebra, black herons and lizards, secretarybirds and crocodiles, a lion pride or two, and our deadliest predators – a leopard and a pack of hyenas.
The south African Highveld, the kind of savannah our ancestors evolved in over four million years ago. Photo: Simon Brown.
What she also saw, and which none of us will ever see, is other groups of human beings that were not H. sapiens. Like our ancestor, they were striding on two legs and using their large brains and opposable thumbs to harvest nuts and berries, sometimes to hunt or scavenge for meat, and to fend off predators. They looked very similar to us, used tools, and some may even have created art and used language to talk to one another.
Around 350,000 years ago, at this stage the earliest date we know H.sapiens might have first strode the planet[i], there was no reason to think things would ever change.
But to say that we are human today is to say that we are members of a single worldwide species. This is extraordinary because for millions of years to be human meant that you could be a member of any one of a number of different but related species.
It should not be contentious to say that all members of the genus Homo are human – after all, this is what the Latin word ‘homo’ means – but it is contentious to suggest, as I will later in these posts, that all bipedal great apes are human.
But first it’s important to state that it’s currently impossible, and may forever be impossible, to finally determine when we stopped simply being hominids – that is, all the apes except for the gibbon – and became hominins as well – that branch of the hominids exclusive to us and our human cousins; this is the point at which chimpanzees went their way and we went ours.[ii] We may never know exactly when the thousands of physical and psychological characteristics that distinguish us from other great apes evolved; what we can be sure about is that almost of them were shared with at least some of our hominin ancestors.
We are so close to our cousins, genetically and historically, that making a distinction between whether or not they are human seems farcical. Indeed, the same argument can be made for any two species close to each other in the hominin line.
In 2005, British celebrity Alan Titchmarsh allowed professional make-up artists to disguise him as a Neanderthal; he then walked along the streets of London, almost completely ignored by everyone.[iii]
Homo sapiens or H. neanderthalensis? Courtesy of Creative Commons. Photographer unknown.
At some point we need to demarcate between those species we consider human and those we consider pre-human, and to date the only specific marker that distinguishes all of us from all of them is bipedalism, not some arbitrarily determined measurement of brain capacity, morphology or dentition.
As well, research into the workings of the human brain, and into animal intelligence generally, has thrown into doubt those psychological characteristics we traditionally considered to be peculiarly human, characteristics that made us special and put us above the rest of the animal kingdom. Once upon a time, we were considered the only animal to make tools, then the only animal to make tools and smile, then the only animal to make tools, smile and do handstands.
It’s similar to a town building a bridge and claiming it’s the only bridge in the world, only to discover that a nearby town has one as well. So the first town now claims it’s the only single-span bridge in the world, until it learns there is a another single-span bridge in the next county. The first town now claims it is the only single-span bridge in the world with green arches, and so on, every new definition increasingly trivialising what makes its bridge special.
There is strong evidence that intelligence has arisen many times in the animal kingdom: in primates, cetaceans, elephants, larger carnivores such as dogs, hyenas and the big cats; birds, particularly corvids and parrots; and some molluscs such as octopuses and possibly squids.
There is also growing evidence that self-awareness and even a theory-of-mind[iv] exists in other primates such as chimpanzees and some birds such as crows.
There is growing evidence that animals other than humans, such as chimpanzees, have a Theory of Mind. Courtesy of Creative Commons. Photographer unknown.
So what are the characteristics that separate humans from our nearest living relatives, the chimpanzee and bonobo?
Before we answer that, we have to talk taxonomy and cladistics – how scientists classify living things.
Life is a spectrum
In his book The Vital Question, biochemist Nick Lane writes that ‘the distinction between a “living planet” – one that is geologically active – and a living cell is only a matter of definition … Here is a living planet giving rise to life, and the two can’t be separated without splitting a continuum.’[v]
Different scientists may employ different markers or waypoints in determining the start of life on Earth, all of which are subject to controversy and disagreement, but the truth is that there is no precise point in time when anyone could claim that a given chemical process for the first time was created by life rather than geology; it would be an arbitrary decision.
The same principle applies throughout evolution. There is no precise point in time where we can say fish gave rise to amphibians or basal reptiles to dinosaurs.
Changes in life brought about by evolution through natural selection isn’t episodic, it’s a spectrum.
But evolution does present a handful of events when with some certainty we can say a new direction had begun – a direction with significant ramifications for all life that follows.
The first of these, and covered in some detail in The Vital Question, concerns the creation of the eukaryotic cell: a morphologically complex cell that contains a separate nucleus and mitochondria, each surrounded by a double membrane. As far as we know this remarkable event occurred only once in all history[vi]. An archaeon, a single-celled prokaryote, absorbed another kind of prokaryote – a bacteria – and instead of consuming it established a symbiotic relationship.
The eukaryotic cell – morphological complexity derived from the synthesis of two prokaryotes, an archaeon and a bacterium. After the creation of life itself, this synthesis is perhaps the most significant event in the history of our planet. Courtesy of Creative Commons.
At some point later in history, some of the descendants of that first complex cell started a symbiotic relationship with a second prokaryote invader, creating chloroplasts and starting the line that would eventually lead to plants and green algae.
More recently, the arrival of the first human was an event with tremendous ramifications for all life on earth.
But when did this happen?
The king of Spain did what?
Before we go any further, we need to talk about a subject that normally works like a sedative on anyone not interested in taxonomic detail: the organisation of the taxa themselves.
I promise to keep this short and to the point, but it’s important to cover because we need to reconsider how and where our human family fits in with other living thing. And, of course, when it all happened.
Mnemonics are as much a part of school science classes as microscopes and Bunsen burners. For example, one mnemonic frequently used in the last century for memorising the different taxonomic ranks was ‘King Philip Came Over From Great Spain’, a mnemonic for the main taxa in the Linnaean system:
Courtesy of Creative Commons.
Kingdom,
Phylum,
Class,
Order,
Family,
Genus, and
Species.
Taxonomic ranking has been around for as long as humans have been curious about the natural world, but the above ranking developed from a system introduced by the Swedish naturalist Carl Linnaeus in the 18th century. He wanted to organise living things so their biological relationship to each other was made very clear. He did this by using shared characteristics to lump things together.
For example, these days all animals with fur, warm blood and that suckle their young with milk are put into one group, the class called mammals. The mammals themselves are grouped together with all animals with a backbone to form a phylum called the chordates. The chordates and animals without a backbone are thrown together into a kingdom called Animalia. More formally, taxa – the collective noun for the rankings – sharing a more recent common ancestor are more closely related than they are to taxa which share a more remote common ancestor: in other words a wombat is more closely related to a dog than it is to a crocodile, and more closely related to a crocodile than it is to a flatworm.
Linnaean taxonomy also introduced the binomial, the familiar two-name identifier used in science to classify an organism at the most detailed commonly used level, that of species. Homo sapiens, for example, is the binomial for human beings, just as Panthera leo is the binomial for lions and Quercus robur is the binomial for the English oak. The first word is the genus (plural genera), the second the species.
Taxonomy is a lovely idea, and appeals to anyone who thinks good old common sense is all you need when sorting bookshelves and tidying kitchen cupboards. For over two hundred years it was regarded as an almost fool-proof system: a place for every living thing and every living thing in its place.
But our knowledge of the natural world is not like that of our kitchen. Like the natural world itself, it is messy, chaotic, growing and constantly evolving.
In 1990, American microbiologist Carl Woese (1928-2012) suggested a new step was needed at the top of the taxonomic ladder to reflect the discovery of a whole branch of life whose existence was never suspected until the 1970s. The archaea, single-celled prokaryotes, were long thought to be a kind of bacteria, but work by Woese and other scientists revealed they are as chemically different from bacteria as we are.
The commonly accepted taxonomic ranks now start with ‘domain’, leaving us with cumbersome and self-defeating mnemonics such as ‘Determined, Kind People Can Often Follow Ghostly Screams’ or ‘Do Kings Prefer Chess On Fridays, Generally Speaking’.
Domain isn’t the only extra rank added over the decades. We also have ‘subfamily’, ‘tribe’, and sometimes ‘subtribe’, ‘subgenus’ and ‘subspecies’, and that’s just in the field of zoology.
In the story of ‘Us’ we’ll be dealing mainly with genus and species, and in the next post we’ll discuss what makes up both taxa.
[ii] Some palaeoanthropologists include chimps and bonobos in the hominin. Rather than outlining all the arguments for or against, I’ll err on the side of caution and include only our immediate family in the hominins.
[iv] This is the ability to attribute mental states similar to your own to other members of at least your own species and possibly other species as well.
[v] Lane, Nick; The Vital Question; London, 2015; p 27.
[vi] This may have happened a second time. A single-celled organism with a nucleus, and possibly mitochondria, dubbed Parakaryon myojinensis, was retrieved a few years ago from the foot of a sea creature found off a coral atoll not far from Japan.
The main difference between P. myojinensis and all other eukaryotes is that its nucleus and mitochondria are surrounded by a single membrane instead of a double one, and its DNA is stored in filaments (as in bacteria) suggesting it is the result of a different line of evolution from all other eukaryotes. Indeed, there is some argument as to whether it is a true eukaryote at all. The only thing that can be said with some certainty is that it is definitely not a prokaryote.
No other example of this creature, or anything similar, has since been recovered. Nonetheless, when it comes to science, hope springs eternal …