Missing something? Perhaps. Earth may once have had rings. Photo: NASA, The Blue Marble.
When I was a wee lad with a growing interest in astronomy and science fiction, I sometimes wondered what it would have been like if Earth, like Saturn, had a ring.
As it turns out, not great, apparently.
In a paper published in Science Direct, Andrew Tomkins, Erin Martin and Peter Cawood suggest Earth may indeed have had a ring during the Ordovician, around 466 million years ago (Ma) with intriguing but catastrophic consequences.
As Tomkins explains in an article for The Conversation, the first clue for the team was an anomalous series of 21 impact craters from around that time, combined with ‘deposits of limestone across Europe, Russia and China containing very high levels of debris from a certain type of meteorite.’
The team then mapped where the craters would have been 466 Ma by using models of tectonic plate movement. Turns out they were all along the equator, with none near the poles.
As Tomkins writes, ‘… we measured how much of Earth’s land surface suitable for preserving a crater was near the equator at that time. Only about 30% of the suitable land was close to the equator, with 70% at higher latitudes.’
So, odds on something peculiar was going on.
The team speculates that Earth captured a passing asteroid which got close enough to exceed the Roche limit (where a smaller body’s internal gravity is overwhelmed by the tidal forces of a larger body). The broken remains of this asteroid then formed a ring around the Earth’s equator, and over time bits of it crashed into the Earth’s surface, forming the impact craters.
Finally, Tomkins et al speculated that the ring would have cast a shadow across large swathes of the Earth’s surface, which together with atmospheric dust from the impacts may explain the sudden plunge into what’s called the Hirnantian Ice Age about 445 Ma.
The team then further speculates that this rapid cooling would have forced animals to adapt to changing conditions, leading to the Great Ordovician Biodiversification Event. Uncomfortably abbreviated to GOBE, this was when Palaeozoic fauna replaced Cambrian fauna, leading to ‘… a spectacular increase in marine biodiversity at all taxonomic levels … ‘[i]
So wearing a ring was not all bad news, I suppose.
[i] Servais, Thomas, et al ‘The Great Ordovician Biodiversification Event (GOBE): The palaeoecological dimension’, ScienceDirect. See here.
From the Wahiba Sands in Oman to penguin-covered ice sheets in Antarctica, notable events in Aiyana Zinkand’s life have been accompanied by one song, a song that no matter the location, the circumstances or the sheer unlikelihood of there being any music at all, inevitably emerges like a cicada in summer.
ABBA’s 1976 Europop hit ‘Dancing Queen’.
Aiyana Zinkand, Vientiane. Photo: Simon Brown
‘The crazy thing?’ Aiyana says. ‘I can’t stand ABBA – I’m more of a Black Sabbath and Cream fan – and even though “Dancing Queen” came out the year after I was born, it is everywhere.’
In 2016 she was on a camping safari in Borneo with friend ‘Irish’ Katie. They stopped at a cinder block camp somewhere on the border between Malaysia and Indonesia. There was no electricity but plenty of rats. The safari staff provided the entertainment in the Orangutan Irish Bar, basically a jungle football pitch decorated with Chinese and American flags.
‘We were sitting there, literally picking off leeches as we drank warm beer, when one of the Malaysian workers picked up a moisture-sodden and out-of-tune guitar and started playing ‘Dancing Queen’.
‘It was hard to pick out exactly what he was singing because he barely spoke English, but I knew it was “Dancing Queen”. No one else figured it out until the final verse.’
#
Aiyana was born in Baltimore, Maryland. Family dynamics were tricky, and she grew up in Maryland, Michigan and Florida.
‘I was a good kid. Stole a police car once, but otherwise I’ve always been on the straight and narrow. Hard working and very sporty.’
Aiyana’s sport of choice was ice hockey.
‘I was a rep player, but because I was a kid couldn’t play professionally. I hoped to pick up a university sports scholarship and go down that line.’
But then, when visiting her father and grandparents in Florida, she was involved in a car accident.
‘I broke ribs, an ankle and knee cap, and dislocated a few vertebrae. There goes my illustrious sporting career.’
#
‘I’ve been travelling all my life, and I’ve heard “Dancing Queen” in a least 30 different countries,’ Aiyana says. ‘That song is everywhere. And everyone knows it.’
The first time she remembers hearing it as the background musical motif to remarkable events in her life was in a karaoke bar in Budapest around 2005.
‘I was working on the thesis for a master’s degree in economics. I got a grant from the Smithsonian exploring postwar reconstruction in Eastern Europe, comparing cities that had concentrated on either cultural or commercial development. But to make enough money to live and travel on I also worked as part-time chaperone for German students on cultural trip through middle Europe: Germany, the Czech Republic, Hungary and Austria.
‘I was meeting up with them in Prague. It was a cold miserable night with a blizzard howling outside. I got to my accommodation about 10.00 pm, resulting in being yelled at by the landlady for being so late. Her husband handed me a huge key ring with lots of keys on it; I didn’t quite get what they were all for because the man was drunk and I could barely understand him.
‘It turned out my room had been given to someone else, and the room I’d been given instead had no bathroom of its own, hence all the keys. Anyway, I didn’t really know any of this at the time and went to what I thought was still my room, opened door to the bathroom … and there was a naked woman there. She screamed. I screamed. We all screamed.’
After things calmed down and they sorted out who had which room, they got talking. The woman’s name was Susanna. She was Spanish and married to a Swiss banker.
‘She was running away from her husband and using his credit cards to spend all his money. The only things she had with her were a red hat box, a fur coat and a crate of Dom Pérignon. Her plan was to live the high life until her husband realised what she was doing and cancelled the cards, whereupon she would divorce him.
‘She had no transport of her own, so we did a lot of driving around together. Then we hit Budapest. More blizzards. We found an underground karaoke bar that played crappy hits from the 70s and 80s. Then it happened. Susanne started singing “Dancing Queen”.’
#
When she finished school, Aiyana moved to California. To keep body and soul together she did 27 different jobs.
‘At some point I found myself doing legal research work for an environmental lawyer. This is about the time things were really starting to happen for long term environmental issues. I was still bartending at night and serving coffee in the morning. But I was good with numbers and eventually worked fulltime with this lawyer. He paid bonuses in the form of college classes in economics, business, math – stuff related to his business. I became his financial forecaster for projects he was handling.
‘I earned a degree in economics in 2005, one class at a time, and became an expert in my field: RFPs – Requests for Proposal – where the state and federal governments put out tenders for work. So I ended up working for both the government and the private sector.
‘The work was intense. At one point I had projects running simultaneously between Sudan, Kuwait, Argentina and Alaska. At the time I was living in Washington, D.C., and had calls coming in every 45 minutes, 24 hours a day.
‘So here I was working with international budgets, forensic accounting, travelling and kind-of-living in Ethiopia, Sudan and Afghanistan while doing work for the UN …
‘And I burnt out.’
#
‘On one trip I was on an Argentinian ship on my way to Antarctica – work to do with base consolidation – and the captain manoeuvred us close to the ice shelf so we could look at some penguins. To set the mood, for whatever reason, he put a song on the intercom.
With friends at Aÿ, France. Photo: Aiyana Zinkand
‘You guessed it. “Dancing Queen”.’
Aiyana stresses that sometimes the song was part of normal, even exuberant, events in her life.
‘I remember being in Aÿ, France, with my best friend, drinking champagne and singing “Dancing Queen” at the top of our lungs. This is just one week after leaving Qatar and one day after hearing the song while watching firemen celebrating the start of summer by leaping naked into a canal during the Boat Festival in Leiden. And I heard it recently at the Garage Bar in Vientiane, looking out at a sunset on the Mekong River.
‘So the song’s associated with good events, not just bizarre ones.’
#
Aiyana says it wasn’t just the sheer amount of work alone that got her down.
‘Increasingly, I wanted to do something that honoured my Quaker religion, rather than dealing among other things with the military and weapon contracts, something that started after 9/11.
‘One of the tipping points was when I was involved in final negotiations for a contract dealing with a Senate oversight committee. We were sequestered in a hotel for the duration, which ended up being about 10 weeks. The day we were released I came out of the air-conditioned hotel smack into a heat wave none of us had any idea was going on. It was 109oF.
‘Living like this in my 20s and 30s … well, my life was disappearing.’
She said she had friends who were international teachers who actually ‘lived’ in the places they worked, who interacted with local people and local culture and history.
‘I was visiting one of these friends who taught in Dakar, Senegal. By that time I was thinking about switching from economics to mathematics, which had really started to fascinate me, and career-switching to teaching. It was New Year’s Eve. Fireworks like I’d never seen before. And, of course, running in the background, “Dancing Queen”.
‘I told my friend about my changing interests, and she immediately shouted out to her boss that I wanted to become an international teacher specialising in mathematics. He told me that with my experience – if I got teacher qualifications – a school like his would snap me up.
‘So that’s what I went and did.’
#
The Ikki Woo Woo Tiki Beach Bar. Photo: Aiyana Zinkand
Every year, Aiyana used to take her dad to Ikki Woo Woo’s Tiki Beach Bar at the Thunderbird Hotel on Treasure Island in Florida, on the coast of the Gulf of Mexico.
‘One time we were there my dad heard “Dancing Queen”. Infuriated, he stood up and shouted: “Fuck Abba!”
‘He then walked off into the sunset, ramrod straight, without his walking cane.’
#
Aiyana did her teaching degree in the US between 2010-2012, ending with her second masters.
‘So I changed my life and became a teacher. I lived and taught in Qatar, and then the Netherlands, and now I teach mathematics to students at the Vientiane International School in Laos.’
#
Wahiba Sands, Oman. Photo: Aiyana Zinkand
Aiyana was holidaying in Oman as a last hurrah before leaving the Middle East to live in Europe, and camping in the desert at Wahiba Sands.
‘I got up early so I could watch the sunrise over the desert – one of my favourite things to do. It was 5.00 am.
‘Then I hear it. What the fuck? In the distance I see a camel herder with five camels and a boombox on one shoulder, and blaring out of the boombox was Abba’s “Dancing Queen”.
In 1977 two American microbiologists discovered a whole new branch of life: the archaea. Single cells without a nucleus, archaea are prokaryotes that for a long time were considered to be bacteria. The work of Carl Woese and George E. Fox, however, revealed that archaea are in some respects closer to eukaryotes (cells with a nucleus, which includes all multicellular life such as you and me) than bacteria.
As a result of their discovery, the traditional taxonomic tree, starting with kingdom and ending with species, was – so to speak – recapped. A new level was placed at the top of the tree – the domain – with three members: bacteria, archaea and eukaryotes.
‘So where do viruses fit in?’ I hear you ask.
Good question. Unfortunately, there isn’t a good answer. The issue is confusing because there is no clear understanding of their evolutionary development, or even if they share a common ancestor. Some may have developed from small, free-floating bits of DNA called plasmids, while others may have evolved from bacteria. It’s generally easier to hedge around the issue of whether or not viruses are actually alive rather than confronting it, and they are often simply called infectious agents or biological entities. The main sticking point is that viruses cannot replicate without infecting a host cell, and are therefore entirely dependent for their existence on ‘biological entities’ that are undoubtedly living. An example of how contentious this argument can be, however, is that the bacteria that causes the STD chlamydia can only exist in a host cell … therefore, should this bacteria be considered technically alive?
Now, to complicate matters even further, a preprint article published in bioRxiv earlier this year announced the discovery of ‘viroid-like colonists’ the authors call ‘Obelisks’, which sounds awfully like the proposed title for a 1950s sf monster movie set somewhere in Death Valley.
However, Obelisks are minute, not monstrous. In fact, they’re small enough to fit inside bacteria. The article’s first sentence describes them as a ‘previously unrecognised class of viroid-like elements that we first identified in human gut metatransciptomic data.’ Basically, they were found in samples of human poo. I think I’d prefer to research giant creatures rampaging through Death Valley, but to each their own.
In size they fit somewhere between viruses and viroids (infections strands of RNA), and are rod-shaped, hence their name. And despite only being discovered recently, they are a lot of them. As the authors report: ‘Large scale searches identified 29,959 Obelisks … with examples from all seven continents and in diverse ecological niches.’
At this point, researchers don’t know if Obelisks are truly alive or not, what they evolved from, or if they are harmful or beneficial to their host organisms. In other words, stay tuned.
At the other end of the scale, a paper by two physicists from University College London suggest that dark energy and dark matter may not exist. To put this in perspective, under the currently most popular model of how the universe works – the lambda-CDM model (or, more simply, ΛCDM) – dark energy and dark matter make up nearly 95% of the universe. It’s like suggesting the theory of evolution through natural selection is fine except for the bit about natural selection … and maybe the bit about evolution.
Authors Jonathan Oppenheim and Andrea Russo ‘… consider a proposed alternative to quantum gravity, in which the spacetime metric is treated as classical, even while matter fields remain quantum.’ Making sense of this is way above my pay grade, but The Guardian’s science correspondent, Hannah Devlin, explains the theory this way: ‘(It) envisages the fabric of space-time as smooth and continuous (classical), but inherently wobbly. The rate at which time flows would randomly fluctuate … space would be haphazardly warped and time would diverge in different patches of the universe.’
If this sounds a bit like Doctor Who trying to explain time to Carey Mulligan’s character in the episode ‘Blink’, it may be because the universe is indeed ‘inherently wobbly’. It’s rather unsettling to think that the Weeping Angels might feel quite at home in Oppenheim and Russo’s universe.
Neither paper has been peer-reviewed at this point, but that hasn’t stopped them garnering media attention and commentary from other scientists. At the very least, ‘Obelisks’ and ‘wobbly spacetime’ have stirred the often lethargic currents of scientific orthodoxy; at best, they demonstrate that all scientific knowledge is provisional.
As FBI Special Agent Dana Scully says, ‘Mulder, the truth is out there.’
(Four year old Maddie was playing with her great-grandfather’s collection of nativity figures. While possessing some familiarity with the original story, she also possesses a vivid and excellent story-telling imagination. This is her version of the nativity as recounted to a spellbound and somewhat gobsmacked audience of great aunties and uncles.)
So, Maddie says …
Joseph, Jesus kidnapper.
Just before guests arrived to see the new baby Jesus, everyone in the manger is sleeping. While they are sleeping, Joseph (looking particularly villainous) kidnaps Jesus and takes him to a dark cave.
The Popcorn Man.
When Jesus’s mother, Doreen, wakes up and can’t find him she knows he’s been kidnapped. She also knows who must had done it, but she needs help to rescue him.
Fortunately for Doreen (and Jesus!), just at that moment the King arrives with the Popcorn Man, and they agree to help rescue Jesus.
Team Doreen ready to spring into action …
(Pause in story here as one of the audience members rudely interjects with a question: ‘Why is he the Popcorn Man?’ Maddie [looking at audience member as if he’s an idiot]: ‘Because he feeds Jesus popcorn!’[i] Well, duh … )
Team Doreen having a well-deserved rest after rescuing Jesus.
Together with the King, the Popcorn Man, an additional visiting king, a small army of shepherds, a flying cow and a flying donkey, Team Doreen rushes to the cave and takes Jesus back from the outnumbered Joseph.
A little while later, Jesus once again safe in his crib in the manger, the King kisses Doreen. The Popcorn Man, the spare king, and the shepherds all join in.
That done, and after a very exciting but tiring nativity, everyone lies down to get some more sleep before cohabiting happily ever after.
THE END
[i] For those of you unfamiliar with the symbology associated with the ancient art of making and selling popcorn, the Popcorn Man is often portrayed as one of the Three Wise Men (or Three Kings) who visit Jesus in the manger, particularly the one carrying a small chest or chests of golden popcorn, often mistaken for gold coins in our more venal times.
The longest ‘age’ in human history is the first, called the Palaeolithic or Old Stone Age. It starts with the oldest stone tools we have found so far, from Lomekwi in Kenya, which are 3.3 million years old. The age ends with the beginning of the Mesolithic or Middle Stone Age in the Middle East about 20,000 years ago.
A recent discovery in Africa, however, suggests we’ve got the title wrong. It shouldn’t be the Old Stone Age, but the Old Wood Age.
Wood does not preserve well, especially compared to bone and stone, so there is little evidence for its use in the palaeoanthropological record, and what evidence there is often inferred by small cuts or nicks.
However, in an article[i] published in Nature in September last year, Barham et al describe the discovery of the first structural use of wood predating the arrival of Homo sapiens by at least 100,000 years. Waterlogged deposits at Kalambo Falls in Zambia preserved ‘two interlocking logs joined transversely by an intentional cut notch.’
The bushwillow logs, as discovered. Photograph: Banham et al, Nature (2023)
The logs, made from bushwillow, were dated by luminescence to be 476,000 years old, give or take 23,000 years.
It’s a remarkable discovery, and confirms that our ancestors (in this case probably H. heidelbergensis[ii]), were as adept at using and shaping wood as they were at using and shaping stone. Given the preponderance of wood in the environment compared to usable stone, and given its comparative softness and adaptability, this doesn’t come as a surprise, but nonetheless it’s nice to have some demonstrably worked specimens.
Wood was almost certainly the first product hominins used to make tools, from scrapers to spears. Indeed, it may have been the dominant raw material for toolmaking throughout most of human history.
So maybe the first human ‘age’ should be referred to as the Old Wood Age, with stone making the occasional guest appearance?
Since April last year, when I published my last blog post, a great deal has happened in the world of palaeoanthropology: claims and counterclaims about ritual burial for Homo naledi, the discovery of the first structural use of wood (predating the arrival of H. sapiens by some 400,000 years!), and genomic evidence of a severe bottleneck in our ancestors’ population about a million years ago which, according to the scientists who uncovered it, almost spelled the end of us.
Goodbye Vientiane …
Before I move on to these topics, however, an apology. The reason I have not written any posts for such a long time is that 2023 was a year of running around and turning about. AJ and I originally intended to return to Australia from Laos at the end of June last year, but for reasons beyond anyone’s control her replacement at her international school was unable to take up the post. AJ agreed to stay on for another six months, so we ended up staying in Laos until the week before Christmas 2023. We spent six months at the start of the year preparing for the move – selling off goods and packing what we could for coming home – only to find ourselves unpreparing, so to speak, and unpacking and buying good back for an extended stay. That was followed by another six months repreparing, repacking and reselling … well, you get the idea.
… hello Wagga Wagga.
AJ and I are now, however, back in Australia, ensconced in Wagga Wagga for the next year or so. After that, fingers crossed, our last move will be to our ‘always’ home in Mollymook on NSW’s south coast for a long and fruitful retirement. Unless something else happens and we end up moving overseas again …
Anyway, future posts – as in near future posts (promise) – will cover those matters palaeoanthropological listed above. As well, I hope to put up an interview with someone who has led an extraordinary life strangely – even bizarrely – intertwined with Abba’s Dancing Queen.
In part 4 of a thematic series of posts called Us, I said this about toolmaking:
‘It is with the application and development of tool usage that the first signs of a distinct ‘human’ culture are found in palaeoanthropology. Whereas chimps and some bird species, like humans, use tools made from plants to gather food or built shelter, humans are the first animals to make stone tools, improving on the original material through knapping.’[i]
Goffin’s cockatoo. Image courtesy of Creative commons.
I short-changed chimps, it seems. Not only do they use tools, they use tool sets: in other words they prepare different tools for different jobs.[ii]
More surprisingly, I now discover that I also short-changed birds. I was pleasantly surprised by the new(ish) information about chimps, but astounded by the news[iii] that at least one species of bird – Goffin’s cockatoo[iv] from Indonesia – also makes and uses different tools for different jobs.
I shouldn’t be astounded, of course. In a much earlier post I wrote about research providing evidence that corvids possessed a Theory of Mind. And as the article in The Conversation points out, an Australian bird – the palm cockatoo – is already known to regularly make drumsticks to beat against hollow trunks during courtship. I suppose it’s not a giant leap from all that to learning that at least one non-avian dinosaur could do with a tool box to keep its implements tidy and dust-free.
It seems that Goffin’s cockatoo actually manufactures three different tools – for wedging, cutting and spooning.
Again, as the article points out, this means the cockatoo’s cognitive skills can be compared directly with a chimps. Importantly, they have ‘… been confirmed as the third species that can not only use tools, but can carry toolsets in anticipation of needing them later on.’
The original research paper leading to the article in The Conversation can be found here in the journal Current Biology.
No, not another instalment in the Starship Troopers media franchise, but an exciting development in the war against the bugs that make us sick … and sometimes kill us.
Alexander Fleming’s discovery of penicillin in 1928 was a turning point in our struggle against bacteria-caused infection. Research carried out by Fleming, and subsequently by Cecil George Paine, Howard Florey and Ernst Chain, marked the start of the systematic production and use of antibiotics, at first in developed countries and later worldwide.
But after eighty years of use, antibacterial resistance is increasingly common. A 2014 report from the World Health Organisation states it is a threat ‘to global public health.’ The report found ‘high rates of resistance … in all WHO regions in common bacteria … ‘[i]
A 2016 review of antimicrobial resistance commissioned by the UK Prime Minister estimated that 700,000 people died each year from resistant infections, and that by 2050 ‘ … 10 million lives a year and a cumulative 100 trillion USD of economic output (could be) at risk … ’[ii]
So it’s kind of surprising that a paper published in February in eBioMedicine with the comparatively catchy title ‘A broad-spectrum synthetic antibiotic that does not evoke bacterial resistance’[iii] might garner some attention in the media. But there’s been hardly any attention at all, if any.
These few lines indicate why the paper may prove to be very important indeed in the future:
‘ … a promising compound, COE2-2hexyl, (exhibits) broad-spectrum antibacterial activity. (It) effectively-treated mice infected with bacteria derived from sepsis patients … including a CRE K. pneumoniae strain resistant to nearly all clinical antibiotics tested. Notably, (it) did not evoke drug resistance in several pathogens tested. (It) has specific effects on multiple membrane-associated functions … that may act together to abrogate bacterial cell viability and the evolution of drug-resistance.’
So not only did it treat bacteria (from sepsis patients) in mice, including a highly resistant strain, it did not evoke resistance and – importantly – might act against the evolution of drug resistance.
Hell, maybe COE2-2hexyl should feature in the next Starship Troopers movie. Sounds like it could take on any bug.
The remains of Homo floresiensis, discovered at Liang Bua on the Indonesian island of Flores in 2003, and of Homo naledi, discovered inside the Rising Star Cave in South Africa’s Cradle of Humankind, have played an important part in helping us understand the diversity and complexity of our hominin past.
Homo floresiensis. Photo courtesy of Creative Commons. Created by ATOR.
H. floresiensis, dubbed ‘The Hobbit’ by the media because of its diminutive size, with a brain capacity of around 380 cm3 and standing around a metre tall, was considered by many scientists to be a deformed or microcephalic H. sapiens. However, strong physical evidence such as humeral torsion[i] and a set of teeth unique among hominins[ii] has pretty well ended the debate about its status as a species in its own right. The main disagreement now, considering the size of its brain, is whether or not it should be included in the genus Homo.
And speaking of small brains …
H. naledi was half again as tall as H. floresiensis – about the same height as a large chimpanzee – and although its cranial capacity (between 460 cm3 and 610 cm3) was considerably bigger than the Hobbit’s, it was still well short of a modern human or any of our immediate cousins such as H. neanderthalensis.
Homo naledi. Photo courtesy of Creative Commons.
As I wrote in a previous post, however, brain size is not necessarily a reliable indicator of intelligence.[iii]H. floresiensis almost certainly made and used stone tools[iv], and recently the University of Witwatersrand’s Lee Berger announced that researchers had found evidence of fire being used by H. naledi[v]. This last was probably something of a given, since the remains of H. naledi were found in a chamber of the Rising Star Cave that could only be reached through a long, dark and twisting route that was difficult and dangerous to follow even with artificial light – without some kind of illumination it would have been virtually impossible. Still, this recent evidence adds weight to the case that this species was capable of making and using fire.
As friend and palaeoanthropologist Debbie Argue asks, however, when and how did H. naledi learn to make fire? Could they possibly have acquired the skill from a contemporary hominin, such as H. sapiens? Or was it the other way around? Or did both species learn the trick from a third hominin group?
We’ll probably never know the answer to this question, but it is fun thinking about, and – at the risk of stretching a metaphor almost to breaking point – throws another log on the fire of revaluating exactly what it means to be human.
[iii] (And in an even earlier post I write about evidence suggesting corvids, with comparatively lightweight brains (c. 20-25 grams, give-or-take), may have a Theory of Mind.)
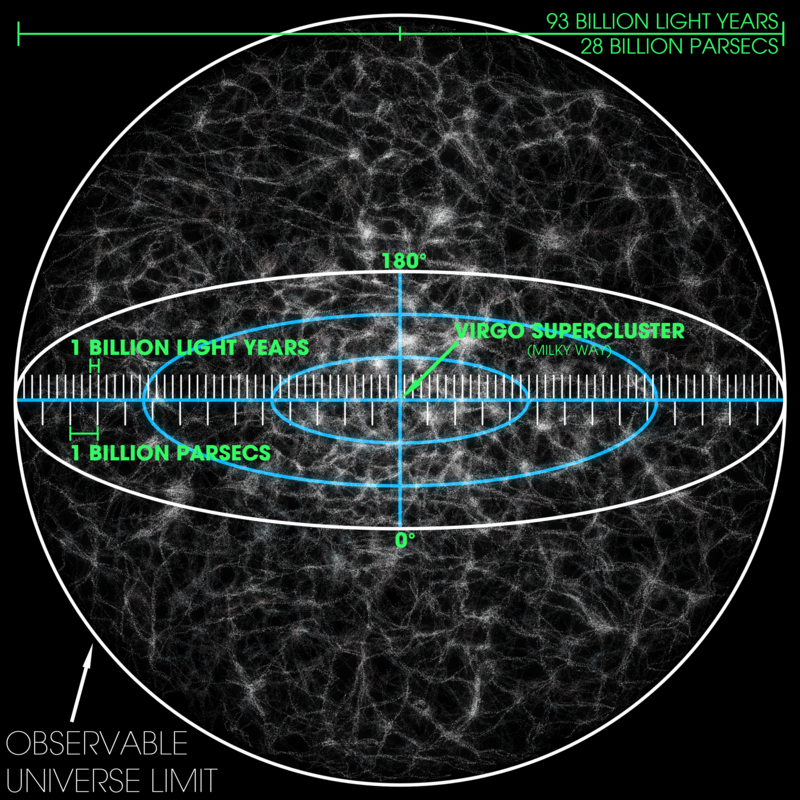
According to a recent article in Quanta Magazine by staff writer Charlie Wood, a recent calculation by two physicists Neil Turok and Latham Boyle, suggests our universe is the most likely option for all universes.
Wood quotes Boyle’s analogy of a sack of marbles, each marble representing a different universe. ‘ … the overwhelming majority of the marbles have just one color — blue, say — corresponding to one type of universe: one broadly like our own, with no appreciable curvature and just a touch of dark energy. Weirder types of cosmos are vanishingly rare.’
The observable universe. Created by Andrew Z. Colvin. Courtesy of Creative Commons.
Turok and Boyle published their calculation in October this year under the extraordinarily catchy heading of ‘Thermodynamic solution of the homogeneity, isotropy and flatness puzzle (and a clue to the cosmological constant)’; but the introductory paragraph contains this killer sentence:
‘The gravitational entropy favors universes like our own which are spatially flat, homogeneous, and isotropic, with a small positive cosmological constant.’
The calculation stems from working with ‘a clock that ticks with imaginary numbers’ enabling Turok and Boyle to calculate the quantity of entropy that corresponds with our universe.
What this might mean for physics is being hotly debated. What is also interesting is the effect it might have on those who think the teleological argument for the existence of god or gods – especially the particular flavour of the argument called the ‘fine-tuned universe’ – has a strong case. This argument states that the universe is special because it is so finely tuned – especially for the existence of life – and that in turn this is evidence of the work of a creator. But if Turok and Boyle are right, then this universe is not so special after all – it is rather common and ordinary. I’m not suggesting this completely negates the argument for a fine-tuned creation, but I think it certainly dilutes it.
However, it is something of a letdown to discover we’re living in a beige universe.