One foot in front of the other
Humans walk upright, gorillas and chimpanzees walk on all fours, resting their weight on their knuckles, and orangutans can do just about anything – they hang and swing by their arms from branches, sometimes with the help of their oddly-shaped feet, and on the ground they can walk either upright or on all fours. The structure of the postcranial skeleton in all four animals is very different and reflects these locomotor patterns. Non-human great apes have short legs and long arms, whereas we have very long legs. With the gorilla and chimpanzee it is the shortness of the legs that differs from humans, the arms being much more similar in length compared to the torso; only the orangutan has enormously lengthened arms. When other great apes stand upright, their legs are straight from hip to ground, whereas humans are ‘knock-kneed’, as the thighs slope inward from the hip to the knee. The pelvis is very different in appearance: in humans the hip bone (ilium) is low and very broad, but in great apes it is high and fairly narrow. In humans the great toe is long and stout and aligned with the other toes, but in great apes it is divergent from the other toes (less in the gorilla), and in the orangutan it is very short.
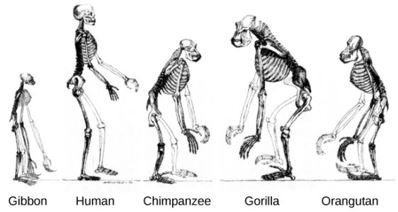
In most great apes, the spinal column is more or less straight, but in humans the spine is curved into a double-S: the cervical (neck) vertebrae curve forward, the thoracic (chest) vertebrae curve backward, the lumbar vertebrae (those in the small of the back) curve forward again, the sacral vertebrae (which are fused together, and form the back wall of the pelvis) curve back again, and the coccyx (the partially fused vertebrae which are the tiny remnant of the tail) curves forward once more. The ribs (which are very variable in number, but average 12 in humans and orangutans, and 13 in chimpanzees and gorillas) together form the thorax; in humans the thorax is barrel-shaped (narrow at the top, broad in the middle, narrower again at the bottom), whereas in great apes it is funnel-shaped (narrow at the top, and broadening towards the bottom).
All of these differences between humans and the other great apes are developments stemming from bipedalism. So why did humans adopt bipedalism? Well, walk with me and we’ll take a brief look at the major theories.
Doing a runner
There seems to be a growing consensus among many scientists that our ancestors evolved bipedalism for several reasons rather than one overriding factor. What many of the competing theories do agree on, however, is that rainforest giving way to savannah because of climate change around 7.0 – 5.0 mya was a strong influence. Grassland with only scattered trees and no closed canopy meant tree-climbing primates had much more open territory to cover. Walking on two legs freed hands to carry infants, food or tools, including weapons. Walking on two legs made us taller, meaning we could locate food, potential predators and safe havens from further away; it also made it easier to pick low-hanging ripe fruit from trees. Walking reduced the amount of body surface area we exposed to the sun while in the open.

Of course, in some circumstances some of these ‘advantages’ could become disadvantages. For example, although bipedalism meant we could locate a predator from further away, it also meant if it was looking in the right direction, a predator could see us from further away as well (and our chief predators – leopards, hyenas and lions – all have good eyesight, not to mention excellent hearing and sense of smell). On the other hand, when our ancestors became active hunters, our extra height gave us an advantage over prey animals, many of whom rely on their sense of smell rather than their eyesight.

More recently, one of the major arguments for the successful adaptation of bipedalism was that it is a much more energy efficient method of locomotion[i]. Whatever the arguments for or against all these hypotheses regarding the origins of walking, when it came to running there is no denying our bodies evolved to make us one of nature’s supreme endurance runners[ii]. This seems to have happened about two million years ago and was a real game-changer when it came to predating: our ancestors evolved into persistence hunters, able to wear down much larger animals such as kudu and oryx[iii]. Basically, humans ran their prey into the ground, and much of our body shape is particularly adapted to long-distance running.
In other words, the characteristics that make us superb walkers and runners are the characteristics that most set us apart from other great apes. As Chris Stringer and Peter Andrews write in The Complete World of Human Evolution, ‘at present … (bipedalism) is taken as the earliest adaptation by which we can recognise human ancestors in the fossil record.’[iv]
The odd-sock drawer
Now it’s time to deal with one of the most controversial species in the human lineage – Homo ergaster. This species was described by Colin Groves and Vratislav Mazák in 1975[v]. Since then, palaeoanthropologists are divided on whether H. ergaster is a distinct species, or a subspecies belonging to H. erectus, palaeoanthropology’s pin-up boy and all-purpose species.
Once they learned to walk, our ancestors just kept on walking. In fact, they walked right out of Africa, into the Middle East, then east into Asia and Sahul, north to Europe, and eventually across the Bering Strait and into the Americas. On the way they continued evolving into new species that seemed to interbreed with each other at every opportunity, creating yet more new species, and eventually discovering agriculture, television and the internet. And interestingly, it’s the use of technology that provides us one piece of evidence that H. ergaster and H. erectus were two different species.
But first, let’s talk more about bones, specifically those belonging to the original H. erectus, parts of which were first discovered 1891 by Eugène Dubois, a Dutch doctor working for the army in Java. In fact, he went to Java with the objective of discovering evidence supporting the theory that H. sapiens evolved in Asia, an idea most determinedly supported by German naturalist Ernst Haeckel. Haeckel had hypothesised that our species’ progenitor, which he names Pithecanthropus alalus, had evolved on Lemuria, a mythical continent that subsequently sunk beneath the Indian Ocean (thereby conveniently leaving no fossils behind to prove – or for that matter, disprove – his theory).

Although Dubois had discovered ancient hominin fossils, he found little or no support among scientists in Europe that they amounted to anything significant. It wasn’t until Sinanthropus pekinensis was discovered in China over a quarter-century later that enthusiasm for Dubois’s discovery really picked up. In the early 1950s, Ernst Mayer reclassified both P. alalus and S. pekinensis as H. erectus[vi]. Since then, hominin fossils with roughly the same estimated brain size as H. erectus and aged between 2 million years old to just over 100,000 years old have been thrown in with H. erectus like differently coloured socks thrown into an odd-sock draw. It has become the species to have when you want to cover all of Africa and Eurasia and two million years of history.
In the early 1970s, for example, Richard Leakey and Alan Walker described two partial skulls found in Kenya as belonging to an African offshoot of H. erectus based on the fact that their calculated brain capacities (848 cc and 803 cc) were not dramatically smaller than that of some H. erectus skulls (around 950 cc), which is like arguing that since the Volvo S60 and the Volkswagen Passat have similar interior space, they’re both examples of a Toyota Camry.
However, in 1975, Colin Groves and Czech colleague Vratislav Mazák, after a comprehensive metric analysis of fossils from Koobi Fora, discovered they had uncovered a new species they names H. ergaster. Their argument was that there was no African version of H. erectus; further, Colin Groves believed that H. ergaster evolved in Africa and then migrated into Eurasia, eventually giving rise to H. erectus.[vii] The earliest dates for the new species goes back 1.9 million years[viii], as opposed to 1.6 million years (or 1.8 according to some estimates) for H. erectus, making H. ergaster the first truly human-looking hominin to stride the planet – tall, thin, decidedly bipedal, with a flatter face than its ancestors and an active hunter, fire-user and tool-maker.

Now, nearly fifty years after the initial paper by Groves and Mazák, a fierce debate still continues between those who think the two hominins are separate if linked species, or just subspecies. In common parlance, it’s a debate between splitters and lumpers.[ix]
But besides the obvious difference in the skull shapes of H. ergaster and H. erectus, another line of evidence convinces me that Colin was right in his opinion that we are talking about two species. This evidence involves tool making.
Out with the old, in with the new
Until the appearance of H. sapiens and H. neanderthalensis, stone age technology is divided into two broad and overlapping stages: Oldowan and Acheulean (sometimes called Modes 1 and 2). Oldowan technology was first discovered in the 1930s by Louis Leakey at the Olduvai Gorge in Tanzania. The oldest examples have been found at Gona in Ethiopia, and date back about 2.5 million years[x]. The technology seems to have spread very quickly, and recent discoveries have found stone tools in Jordan dated at 2.5 mya and China at 2.1 mya[xi]. This technology, the use of very simple flakes and rocks, had been developed before the appearance of H. habilis, possibly by Australopithecus garhi. Acheulean technology which started about 1.76 mya, is closely associated with the appearance of H. ergaster and involves more refined knapping and the development of specialised tools such as hand axes.
This doesn’t imply that Oldowan technology suddenly evaporated, and every hominin adopted the new style of knapping chert. In some places, Oldowan and Acheulean stone tools are found at the same site from the same period, suggesting that while H. ergaster or one of its descendants employed the improved technology, one of our cousins continued using the older method. But it’s clear Acheulean technology obviously conferred a significant advantage over the old style. It didn’t take long for it to spread beyond Africa, either because H. ergaster itself started spreading beyond Africa, or because it spread by ‘word-of-mouth’: neighbouring hominis picked up on the new fashion of making tools and copied it. Acheulean tools appear in what is now India, for example, by 1.5 mya, and in Europe by about 900 kya.

However, Acheulean technology did not seem to reach Java, where our friend H. erectus resided.
Which presents lumpers with a problem. If H. ergaster is indeed nothing more than a subspecies of H. erectus, then fossil evidence suggests this single species arose in Africa before spreading throughout Eurasia. Yet if this is also the species that developed Acheulean technology soon after evolving, why didn’t the technology travel with them to the far east?
On the other hand, if we are talking about two species, then it’s quite possible for Acheulean technology to be developed by H. ergaster in Africa, spread slowly throughout Eurasia, but never quite reach the home of H. erectus in Java.
If this was in fact the case, it raises a more important question: even if we accept H. ergaster is a separate and earlier species than H. erectus. Does it necessarily follow that H. ergaster gave rise to H. erectus? What if the two species are cousins rather than mother and daughter?
This is something we’ll discuss in the next, and final, post of ‘Us’.
Other posts in this series can be found here:
‘Us’ Part 3 – The devil in the detail
‘Us’ Part 4 – Using your noggin
[i] https://www.newscientist.com/article/dn12269-walking-on-two-feet-was-an-energy-saving-step/
But then again, see https://www.sciencedirect.com/science/article/abs/pii/S0047248412001443
[ii] https://www.nature.com/articles/nature03052
[iii] https://en.wikipedia.org/wiki/Persistence_hunting
[iv] Stringer, C. & Andrews, P. The Complete World of Human Evolution. London, 2011. P 19.
[v] https://www.irmng.org/aphia.php?p=taxdetails&id=10031853
[vi] According to Britannica, Mayr did this in 1944. But see Bernard Wood who writes it was Franz Weidenreich who first came up with the idea in 1940:
‘(He) was the first to suggest that the genus Pithecanthropus should be subsumed into Homo, and in the same paper he proposed that fossils recovered from what was then called Choukoutien (now called Zhoukoudian), which were initially assigned to Sinanthropus pekinensis,26 should also be transferred to H. erectus.’
[vii] From the Australian Museum:
‘A growing number of scientists have redefined the species Homo erectus so that it now contains only east Asian fossils. Many of the older African fossils formerly known as Homo erectus have now been placed into a separate species, Homo ergaster and this species is considered to be ancestral to Homo erectus. The redefined Homo erectus is now generally believed to be a side branch on our family tree whereas Homo ergaster is now viewed as one of our direct ancestors. ‘
[viii] Oldest fossil dates according to the Australian Museum for H. ergaster here and for H. erectus here. Recent work reported in the journal Science may push the dates even further back, between 1.95-2.04 mya (although in this paper the discussed specimen is describe as preserving ‘characters that align it morphologically with H. erectus sensu lato (including Homo ergaster)’. Go figure.
[ix] For a fuller description of the often heated debate about what makes a species, see here.
[x] Stringer, C. & Andrews, P. The Complete World of Human Evolution. London, 2011. P 208.
A new kind of stone age technology – Lomekwian – has been suggested after the recent discovery of stone tools at Lomekwi that predates Oldowan by more than 700,000 years. See the previous post for more details.
[xi] https://onlinelibrary.wiley.com/doi/abs/10.1002/evan.21863

6 comments